The frontal lobe is the most rostral and largest lobe in each cerebral hemisphere. It extends from the frontal pole anteriorly to the central sulcus posteriorly. It extends upto the interhemispheric fissure medially and the sylvian fissure laterally. The anatomy of the frontal lobe is best understood by studying its three surfaces viz: the superolateral surface, the medial surface and the basal surface.
Superolateral Surface of the Frontal Lobe

The superolateral surface of the frontal lobe is divided by two longitudinal sulci and one horizontal sulcus into the superior, middle and inferior frontal gyri and the precentral gyrus. The sulci are named as the superior (SFS) and inferior frontal sulci (IFS) and the precentral sulcus (PreCS)respectively. The superior frontal gyrus (SFG) lies between the superior frontal sulcus (SFS) and the precentral sulcus (PreCS). A portion of the superior frontal gyrus extends to the medial surface of the hemisphere. This is usually separated from the dorsolateral portion by a medial frontal sulcus (MFS). This portion of the superior frontal gyrus is also called as the medial frontal gyrus (MeFG). The superior frontal gyrus is continuous anteriorly with the gyrus rectus (GR) and connects posteriorly with the precentral gyrus (PreCG). The most posterior and medial portion of the superior frontal gyrus just anterior to the precentral gyrus is known as the supplementary motor area (SMA).[1]. The area just anterior to the SMA is the pre-SMA. Both the regions have poorly defined boundaries and maybe best identified by intra-operative brain mapping or magnetoencephalography. Both these regions are related to performing learned sequences of movements. The most caudal portion of the superior frontal gyrus that lies on the supero-lateral surface in front of the precentral gyrus is the dorsal premotor area, which is involved in the planning of movements.

The middle frontal gyrus (MFG) is the largest of the three frontal gyri and lies between the superior and inferior frontal sulci. It frequently connects with both the superior and inferior frontal gyri (IFG) and the precentral gyrus (PreCG) through discontinuations or interruptions in the sulci concerned. The most caudal portion of the middle frontal gyrus anterior to the precentral gyrus harbors the frontal eye fields responsible for saccadic and voluntary eye movements.[1]
The inferior frontal gyrus is divided into the pars orbitalis (PO), pars triangularis (PT) and the pars opercularis (pOp) and lies between the inferior frontal sulcus medially and the sylvian fissure (Sf) laterally. The sylvian fissure is divided into an anterior and posterior part by the anterior sylvian point. The anterior sylvian point corresponds to an enlargement of the sylvian fissure recognized intra-operatively as a CSF filled area which lies just beneath the pars triangularis of the inferior frontal gyrus.
From the superior aspect of the anterior sylvian point emerge the horizontal and ascending rami of the sylvian fissure which demarcate the pars triangularis separating it from the pars orbitalis anteriorly and the pars opercularis posteriorly. The pars orbitalis is the anterior most part of the inferior frontal gyrus and connects anteriorly with the orbital gyri. The pars opercularis is U shaped due to the inferior termination of the precentral sulcus.[1] The pars opercularis connects prominently with the precentral gyrus just above the sylvian fissure. In the dominant hemisphere the pars opercularis and the pars triangularis correspond to the Broca’s area, which is responsible for motor speech production. The most posterior portion of the pars opercularis near its connection with the precentral gyrus corresponds to the ventral premotor area and intra-operative stimulation of this area causes speech arrest.[1]



Precentral Gyrus
The precentral gyrus (PreCG) lies between the precentral sulcus (PreCS) and the central sulcus (CS). The central sulcus separates the precentral gyrus from the post-central gyrus posteriorly.
The precentral gyrus corresponds to the primary motor area (PMA) and the postcentral gyrus (PoCG) corresponds to the primary sensory area (PSA). Both the precentral and postcentral gyri are connected to each other at the medial and lateral ends of the central sulcus. The medial connection is the paracentral lobule (PaCL), which lies on the medial aspect of the hemisphere in the interhemispheric fissure. The lateral connection, which lies above or embedded within the sylvian fissure is the subcentral gyrus (SubCG). The central sulcus has been traditionally viewed as having two anteriorly convex genus and one middle posteriorly convex genu. Broca described a third middle connection between these two gyri, which lies in the depths of the central suclus as a gyral bridge also known as the “plis de passage” moyen of Broca.[1] This connection corresponds topographically to the middle posterior convex genu of the central sulcus. This middle knee also corresponds to the motor hand area and can be identified at the posterior end of the superior frontal sulcus where it reaches the precentral sulcus.
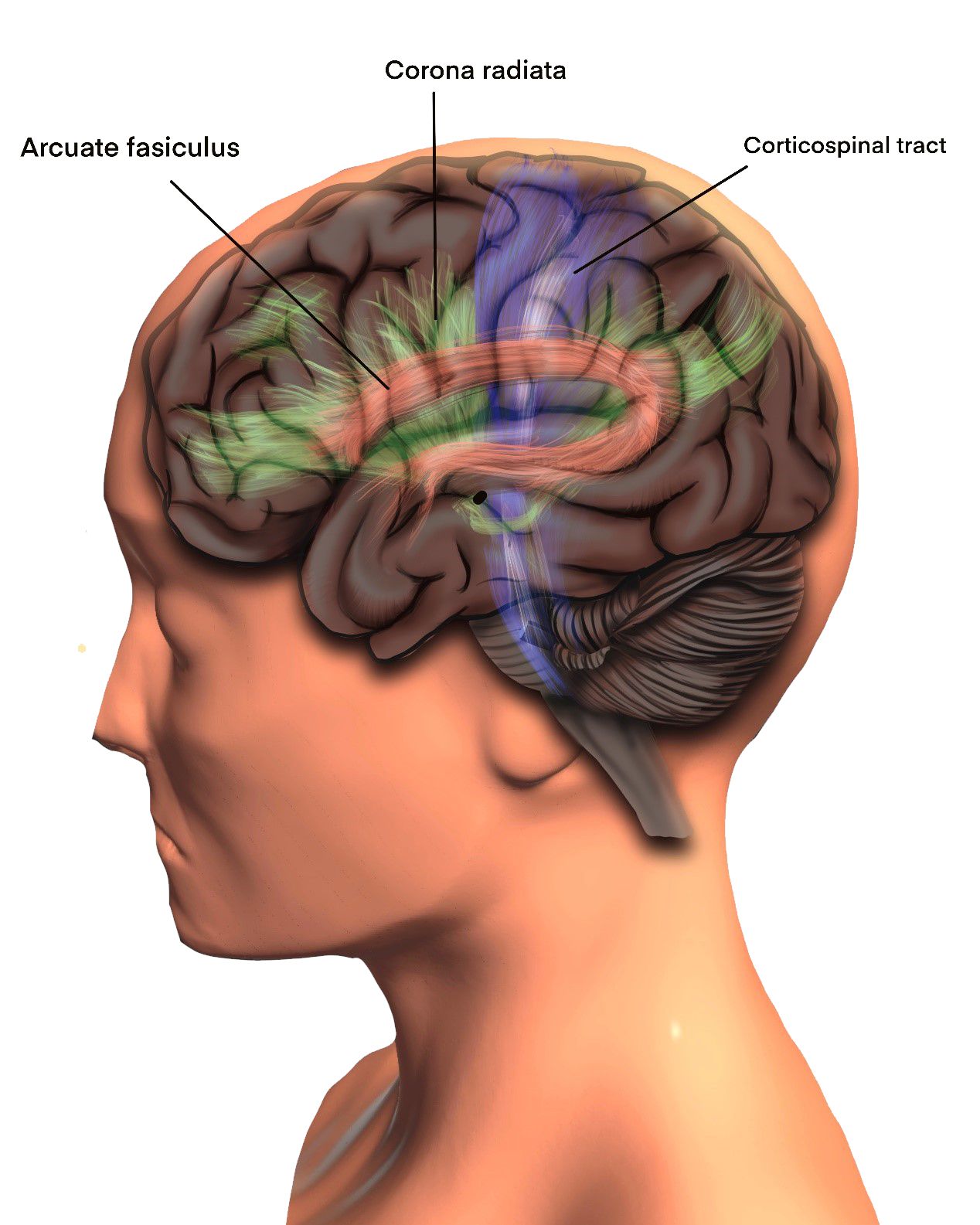
After removal of the superficial and intersulcal grey matter and the short arcuate fibers the long association fibers in the region of the frontal lobe come into view. Beneath the superior frontal gyrus run the fibers of the corona radiata, the callosal fibers and the medial superior longitudinal fasciculus (SLF-I) from lateral to medial.[2] The fibers of the corona radiata are vertically oriented and those of the superior longitudinal fasciculus are horizontally oriented. Medially, deep to the superior longitudinal fasciculus lies the cingulum and deeper still are the transversely running fibers of the corpus callosum. The fibers deep to the middle frontal gyrus are the corona radiata, the superior longitudinal fasciculus (SLF-II) and the arcuate fasciculus from medial to lateral.[2] The arcuate fasciculus (AF) lies ventrolateral to the superior longitudinal fasciculus in this region. Removal of the inferior frontal gyrus exposes the superior longitudinal fasciculus (SLF-III) medially and superiorly and the arcuate fasciculus laterally and inferiorly. Deep to these fibers lie the fibers of the external capsule joining the internal capsule to form the corona radiata that splits the SLF into its medial and lateral portions.[2]
Connections between the superior and middle frontal gyri with the pre-central gyrus are revealed on removal of the superficial gray matter from the latter gyrus. Removal of the short arcuate fibers reveals from medial to lateral the fibers of the superior longitudinal fasciculus (medial part), the corona radiata, the continuation of the superior longitudinal fasciculus (lateral part) and lastly the arcuate fasciculus. [2] [3] The fibers of the arcuate fasciculus curve downwards just posterior to the post central gyrus in the region of the inferior parietal lobule.
Medial Surface of Frontal Lobe
The medial surface of the frontal lobe is composed of the continuation of the medial portion of the superior frontal gyrus superiorly and beneath it the cingulate gyrus (CG). Anteriorly at the frontal pole the orbital gyri meet the superior, middle and inferior frontal gyri. The most infero-medial gyri are the paraolfactory, paraterminal and the subcallosal gyri above which is the beginning of the cingulum ventral to the rostrum of the corpus callosum. [2] The cingulum then runs upwards and posteriorly beneath the gray matter of the cingulate gyrus in the medial portion of the fronto-parietal lobe. Removal of the short arcuate fibers of the frontal and parietal lobe shows the connections of the cingulum to the fronto-parietal cortex. Removal of the short arcuate fibers of the superior frontal gyrus reveals the superior longitudinal fasciculus, the upward curving callosal fibers and the corona radiata from medial to lateral.[2] Beneath the superior longitudinal fasciculus runs the cingulum and beneath the cingulum the transverse fibers of the corpus callosum can be visualized.


Subcortical Anatomy
Superior Longitudinal Fasciculus (SLF)
The superior longitudinal fasciculus is the widest and most superficial of the long association fibers and runs between the frontal and parietal lobes. Traditionally the superior longitudinal fasciculus and the arcuate fasciculus were considered as part of a single fiber system and described as a single structure with a horizontal frontoparietal segment, a vertical temporo-parietal segment and a horizontal frontotemporal or arcuate fasciculus. A few other studies described the superior longitudinal fasciculus as having three parts: SLF- I, SLF – II and SLF – III from medial to lateral. Our dissections have revealed the superior longitudinal fasciculus to be a broad sheet of fibers with wavy layers running from the frontal to the parietal regions of the brain.[2] [3] This sheet is divided by the projection fibers into medial and lateral parts. The medial part of the SLF (also known as the SLF – I) begins at the superior frontal gyrus and runs posteriorly to reach the precuneus in the parietal lobe. It runs above the cingulum and its medial relations are the upcurving fibers of the corpus callosum and the corona radiata. The lateral part of the superior longitudinal fasciculus (SLF-II and SLF – III) lies at a lower level than the medial portion due to the configuration of the cerebral convexity. This part of the SLF connects the middle frontal gyrus to the angular gyrus (SLF – II) and the inferior frontal gyrus to the supramarginal gyrus (SLF – III). It is not very easy to separate these two portions of the SLF that are commonly seen as lying one on top of the other as they run between the frontal and parietal lobes. The arcuate fasciculus lies ventral to this portion of the superior longitudinal fasciculus. The mean length of the superior longitudinal fasciculus was 13.5 cm (12.2 to 14.7 cm). It connects widely various regions of the frontal, parietal and temporal lobes. The fasciculus lies at a depth of 2 to 2.5 cms from the surface.
Function:
The SLF I connects the motor and pre-motor cortices to the superior parietal cortex whose function is encoding location of body parts in a body-centric coordinate system.[2][3] Thus it seems that the medial part of the SLF (SLF – I) is involved in regulating motor behavior especially conditional associative tasks, which select among competing motor tasks based on conditional rules. The SLF II connects the dorsal prefrontal cortex to the caudal inferior parietal cortex, which controls spatial attention and visual awareness. This suggests the SLF II provides the prefrontal cortex with parietal cortex information regarding perception of visual space. Since these bundles are bi-directional, working memory in the prefrontal cortex may provide the parietal cortex with information to focus spatial attention and regulate selection and retrieval of spatial information. The SLF III connects the supramarginal gyrus to the ventral premotor and prefrontal cortex. This suggests that the SLF III transfers somatosensory information, such as language articulation, between the ventral premotor cortex, pars opercularis, the supramarginal gyrus, and the lateral inferior prefrontal cortex. In the dominant hemisphere this pathway is important for the articulatory aspects of language and in the non-dominant hemisphere it subserves visuospatial attention, prosody and music processing. [2]
Arcuate Fasciculus
The arcuate fasciculus (AF) is a reverse C-shaped fiber bundle that connects the Broca’s and the Wernicke’s area.[2] [4] It is described as having a dorsal and a ventral segment. In our dissections, the AF was seen running inferior to the lateral portion of the SLF in the frontal and parietal regions, then curving downwards in the region of the supramarginal and angular gyri and ultimately running forwards towards the temporal pole beneath the superior and middle temporal gyri. The length of the arcuate fasciculus was 12.5 cm2. Thus the AF wraps around the posterior edge of the insula in a horseshoe-like configuration; connects the middle, posterior, and anterior parts of the middle and superior temporal gyri to the middle parts of the inferior and middle frontal gyri; and passes deep to the middle and superior part of the lateral surface of the temporal lobe, anterior part of the occipital lobe, angular and supramarginal gyri, middle and inferior parts of the pre- and postcentral gyri, and anterior and middle parts of the middle and inferior frontal gyri.[2] [4] Functionally, evidence for the role of the AF in language use is best represented by conduction aphasia. This type of aphasia manifests as an inability to repeat unfamiliar sounds.
The classical model of language consists of the Broca’s area, Wernicke’s area and the AF connecting these two regions. Recently Hickok et al proposed a dual stream model for language, the dorsal stream and the ventral stream.5 The dorsal stream is involved with phonological processing and is mediated by the lateral portion of the superior longitudinal fasciculus (SLF II and III) and the AF. The ventral stream of language is involved with semantic processing and consists of the inferior fronto-occipital fasciculus (IFOF), the inferior longitudinal fasciculus (ILF) and the middle longitudinal fasciculus (MLF).
The cingulum is a C-shaped association bundle which runs superior and parallel to the corpus callosum. Its fibers terminate anteriorly below the rostrum of the corpus callosum in the subcallosal gyrus.4 Posteriorly the cingulum narrows above the splenium, where it forms the isthmus of the cingulum and this continues below as the radiation of the cingulum to end in the anterior parahippocampal gyrus, adjacent to the hippocampus. The length of the whole cingulum from its beginning in the subcallosal area to its termination in the parahippocampal gyrus was found to be 19 cm. The cingulum lies at a depth of 3.5 cm from the convexity surface of the brain. The cingulum, from its superior surface, sends connections to the frontal and parietal cortices. Functionally, the cingulum is a major component of the dorsal limbic pathway. Damage to this structure leads to behavioral and emotional disturbances. It is also a central structure in learning to correct mistakes and is involved in the appraisal of pain and reinforcement of behavior that reduces pain.
Corona Radiata
The external and internal capsule fibers join together at the upper edge of the putamen to form the corona radiata (CR).[2] [6] [7]The CR runs through majority of the frontal and parietal lobes and bisects the middle group of association fibers into medial and lateral halves. At its medial margin, the callosal fibers join the CR to form the centrum semiovale. Functionally, damage to the CR can cause varying degrees of contralateral motor and sensory deficits.
Cortical Anatomy
The orbital surface of the frontal lobe is divided into the medial gyrus rectus and the lateral orbital gyri by the deep olfactory sulcus.[2] The olfactory sulcus houses the olfactory bulb and the olfactory tract. The orbital gyri make up most of the basal portion of the frontal lobe. An H-shaped sulcus, also known as the cruciform sulcus of Rolando, divides the orbital gyri into anterior, posterior, medial and lateral orbital gyri. The posterior orbital gyrus is situated just anterior to the anterior perforated substance and connects medially to the medial orbital gyrus. The medial, anterior and lateral orbital gyri are connected to the superior, middle and inferior frontal gyri along the frontal pole.
The olfactory tract runs in the olfactory sulcus on the orbital surface of the frontal lobe. Further posteriorly, it divides into the medial and lateral olfactory striae. The medial olfactory stria becomes continuous with the subcallosal and the paraterminal gyri on the medial surface of the frontal lobe. The lateral olfactory stria passes along the lateral and anterior margin of the anterior perforated substance, hugging the posterior margin of the posterior orbital gyrus, to reach the piriform region in the temporal lobe. The medial stria contains fibers from the anterior olfactory nucleus. These fibers traverse in the anterior commissure (AC) to the opposite side. The lateral olfactory stria passes along the lateral margin of the anterior perforated substance, giving out some collaterals into the anterior perforated substance, to reach the piriform region. These fibers terminate in the prepiriform cortex and the corticomedial group of the amygdaloid nuclear complex. The prepiriform area represents the primary olfactory cortex. It corresponds to area 34 of Brodmann and is located on the anterior end of the parahippocampal gyrus and uncus. From here fibers project to the neighboring entorhinal cortex and the medial dorsal nucleus of the thalamus. Fibers from the amygdaloid nuclei connect with the hypothalamus and the septal nuclei.

Subcortical Anatomy
The first fibers that are visualized after removing the superficial gray matter of the orbital surface of the frontal brain are the arcuate fibers. These are short association fibers that interconnect neighboring gyri. Removing the short association fibers in the region of the medial gyrus rectus will reveal the anteriorly curving fibers of the forceps minor. Inferior and lateral to the fibers of the forceps minor are association fiber bundles that connect the orbito-frontal gyri to the supero-medial portion of the thalamus. These fibers run lateral to the gyrus rectus, inferior to the frontal horn and further posteriorly run lateral to the caudate nucleus. Deeper dissection in the region of the medial orbital gyrus will reach the frontal horn. The caudate head lies lateral to the frontal horn and roughly corresponds to the transverse limb of the H-shaped cruciform sulcus. Fiber bundles running lateral to the caudate connect the orbitofrontal gyri to the midbrain via the crus cerebri. The medial portion of the orbitofrontal region thus connects to the thalamus and the brainstem. The lateral portion of the orbitofrontal gyri mainly connects with the ipsilateral temporal, parietal and occipital cortices by two major association bundles, namely the inferior fronto-occipital fasciculus (IFOF) and the uncinate fasciculus.
Inferior Fronto-Occipital Fasciculus (IFOF)
Fibers from the lateral orbital gyri join fibers from the middle and inferior frontal gyri to form the IFOF.[2][8][9][10] All the fibers aggregate and pass lateral and superior to the corona radiata fibers, and medial and inferior to the superior longitudinal fasciculus (SLF) and arcuate fasciculus towards the temporal lobe, as a constituent of the temporal stem. In the region of the insula, the IFOF and the uncinate fasciculus form the ventral portion of the external capsule, with the IFOF lying superiorly and the uncinate fasciculus lying inferiorly. After exiting the temporal stem the majority of the IFOF passes through the superior and middle temporal gyri to reach the parietal and occipital gyri. Some of its fibers turn anteriorly along with the uncinate fasciculus to end in the superior and middle temporal gyri. The fibers of the IFOF running posteriorly form a component of the sagittal stratum and lie lateral to the optic radiations (OR). Fibers running medially in the IFOF project to the parietal lobe and fibers running laterally project to the occipital lobe. Functionally, the IFOF forms the main portion of the ventral semantic pathway. It is believed to be involved in semantic processing, reading, writing, comprehension and production of meaningful speech.[2] It is also involved in visual recognition, integration of multimodal sensory input and motor planning. Intra-operative stimulation of the IFOF gives rise to semantic paraphasias (error in the meaning of the target word).
Uncinate Fasciculus
The uncinate fasciculus (UF) is a hook shaped tract running in the temporal stem connecting the orbitofrontal and temporal regions.[2][6][7][8][9][10] It originates in the orbitofrontal gyri, runs laterally deep to the IFOF and anteroinferior to it in the temporal stem, and ends in the superior and middle temporal gyri. The dorsolateral portion of the UF connects the temporal pole to the lateral orbitofrontal gyri and its ventromedial portion connects the temporal pole to the medial orbitofrontal cortex and the septal area. The frontal portion of the UF courses just anterior to the anterior perforated substance. It covers the inferior and medial sides of the nucleus accumbens and reaches the region below the genu of the corpus callosum. Functionally, the UF is considered to be a part of the ventral limbic pathway.[2][6][7][8][9][10] The literature refers to three functions of the UF: associative and episodic memory functions, linguistic functions and social–emotional functions.[2] UF involvement is seen in five disorders, namely, anxiety, schizophrenia, psychopathy, epilepsy and frontotemporal dementia
Anterior Commissure
The anterior commissure (AC) crosses the midline anterior to the columns of the fornix.[2] [4] [8] [10] The entire extent of the AC resembles a horizontally placed bow. It runs in a canal of gray matter (canal of Gratiolet) parallel and deep to the UF and the occipitofrontal fasciculus. The average distance of the AC from the temporal pole is 18 mm (16 to 20 mm) and the average distance of the UF from the AC is 6 mm (3 to 8 mm).[2][4][10] Substantia innominata lies in the depth between the UF and the AC. Laterally, the AC splits into two, an anterior or olfactory portion and a posterior or temporal portion. The anterior portion consists of fibers arising from the anterior olfactory nucleus, cross to the opposite side and end in the anterior olfactory nucleus on the contralateral side. The posterior or the temporal portion forms the major bulk and passes laterally, deep to the lentiform nucleus, to end in the middle temporal gyrus. Some fibers enter the external capsule laterally and some fibers course posteriorly as part of the sagittal stratum. Functionally the AC plays a role in the transfer of visual, olfactory and auditory information between the temporal lobes.
Basal Forebrain
The mediobasal frontal cortical area of each cerebral hemisphere, composed of the paraterminal gyrus and the paraolfactory gyrus, is considered a limbic cortical area.
It extends from the olfactory tubercle rostrally to the hypothalamus caudally. It lies over the anterior perforated substance and does not have any definite boundaries. The basal forebrain includes the septal area, the olfactory tubercle, parts of the amygdala and the substantia innominata. The term substantia innominata is also used synonymously with the term basal nucleus. The lateral olfactory stria passes along the lateral margin of the anterior perforated substance to reach the piriform region. These fibers terminate in the piriform cortex and in the corticomedial group of the amygdaloid nuclear complex.
The medial olfactory stria becomes continuous with the subcallosal and the paraterminal gyrus. The subcallosal area and the paraterminal gyrus constitute the septal area, beneath which are the septal nuclei. The septal region is situated on the medial surface of the cerebral hemispheres, immediately facing the AC. This region is also known as the septum verum (“true septum”), or precommissural septum, as opposed to the septum pellucidum, which does not contain neuronal cells and is situated posterior and superior to the AC, morphologically constituting the medial walls of the frontal horns and the ventricular bodies. The medial septal nucleus becomes continuous with the nucleus and tract of the diagonal band of Broca, which then connects with the hippocampal formation.
The septal nuclei receive afferents principally from the hippocampus and subiculum of the parahippocampal gyrus via the indusium griseum, as well as from the precommissural fornix. The efferents of the septal nuclei project to the area surrounding the hippocampus via the fornix itself, to the hypothalamus and midbrain tegementum via the medial forebrain bundle, to the habenula via the stria medullaris thalami, which in turn projects to the mesencephalon via the fasciolus retroflexus. Functionally, the septal nuclei are responsible for connecting limbic structures with the hypothalamus and the brainstem, principally via the hippocampal formation. The so-called “septal syndrome” is clinically characterized by exaggerated reactions to environmental stimuli and the consequent behavioral alterations, principally related to eating and drinking habits, as well as by episodes of rage and disorders in the sexual sphere. Afferents to the basal forebrain arise from the amygdala, the insula, temporal, piriform and entorhinal cortices. From the basal forebrain, neurons project to various regions of the cerebral cortex.
REFERENCES
1. Ribas GC. The cerebral sulci and gyri. Neurosurg Focus 2010; 28(2):E2.
9. Rhoton AL. The cerebrum. Neurosurgery 2007; 61(Supp 1):37–119.